Vetzuurstofwisseling
Vet verbranding (Vet afbraak)
Omdat de vetverbranding een ingewikkeld proces is, is dit onderdeel verdeeld in een aantal verschillende onderdelen. Hieronder staan de verschillende onderdelen vermeld.
Triglyceriden worden gehydrolyseerd door cyclisch AMP-gereguleerde lipasen
De eerste gebeurtenis in het gebruik van vet als energiebron is de hydrolyse (= afbreken m.b.v. water) van triglyceriden door de enzymen die lipasen worden genoemd. Dit proces wordt ook wel lipolyse genoemd. Lipasen zetten triglyceriden om in glycerol en vetzuren, zie hiervoor de onderstaande figuur.
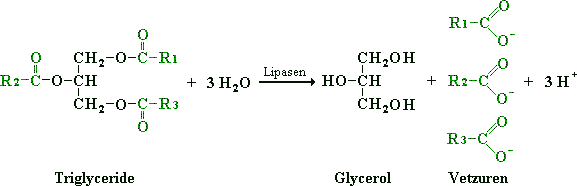
Figuur: De hydrolyse door lipasen van triglycerol in glycerol en vetzuren.
De activiteit van lipase in vetcellen wordt gereguleerd door hormonen zoals adrenaline en glucagon. Deze hormonen activeren het enzym adenylaatcyclase. Dit enzym zet ATP om in cyclisch AMP. Dit cyclisch AMP activeert op zijn beurt weer het enzym proteine kinase A (PKA). Het enzym PKA fosforileerd het lipase en wordt hierdoor geactiveerd. Net zoals bij de afbraak van glycogeen is cyclisch AMP hier "de second messenger". Het hormoon insuline remt de hydrolyse van triglyceriden.
Glycerol, dat door de afbraak van triglyceride is ontstaan, wordt gefosforileerd door glycerolkinase en vervolgens geoxideerd door glycerolfosfaatdehydrogenase tot dihydroxyacetonfosfaat. Dit is een intermediair van de glycolyse en zal hierin verder worden afgebroken.
Vetzuren worden gebonden aan coenzym A voordat zij worden geoxideerd
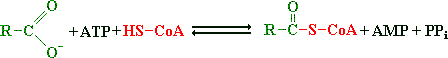
Figuur: Een vetzuur reageert met ATP en coenzym A tot acyl CoA, AMP en pyrofosfaat.
Een vetzuur reageert met ATP en coenzym A tot acyl CoA, AMP en pyrofosfaat. Deze reactie wordt gekatalyseerd door acyl CoA synthetase.
Het enzym acyl CoA synthetase is gebonden aan het buitenmembraan van het mitochondrium. De totaalreactie ligt sterk in de richting van acyl CoA door de snelle hydrolyse van pyrofosfaat (een terugkerend patroon in de biochemie).
Carnitine transporteert lange-keten geactiveerde vetzuren de mitochondriële matrix in
Vetzuren worden geactiveerd aan de buitenmembraan van het mitochondrium, maar worden geoxideerd in het mitochondrium. Omdat lange-keten vetzuren niet gemakkelijk door de binnenmembraan van het mitochondrium gaan is hiervoor een speciaal transportmechanisme nodig.
Geactiveerde lange-keten vetzuren worden samengevoegd met carnitine. De acylgroep wordt van het zwavelatoom van coenzym A overgedragen op de hydroxylgroep van carnitine onder vorming van acylcarnitine. Deze reactie wordt gekatalyseerd door carnitineacyltransferase I, dat is gebonden aan de buitenmembraan van het mitochondrium.
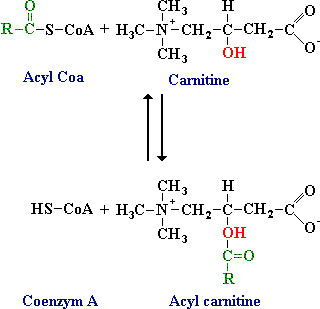
Figuur: Geactiveerde lange-keten vetzuren worden samengevoegd met carnitine.
Acylcarnitine wordt dan door het binnenmembraan verplaatst door een translocase enzym (membraaneiwit). De acylgroep wordt terug overgedragen aan coenzym A aan de matrixkant (in het mitochondrium) van het membraan. Deze reactie die wordt gekatalyseerd door carnitine acyltransferase II. Tenslotte wordt carnitine naar de cytoplasmatische kant teruggetransporteerd door het translocase in ruil voor een binnenkomend acylcarnitine.
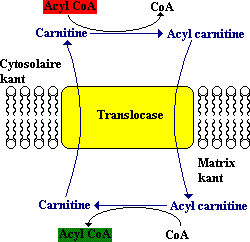
Figuur: De verplaatsing van acryl carnitine in de mitochdrische matrix wordt gekatalyseerd door translocase.
Vetzuren worden afgebroken door afsplitsingen van steeds twee koolstofatomen
Vetzuren worden afgebroken door op elkaar volgende afsplitsingen van delen met twee koolstofatomen. De reacties die elkaar opvolgen zijn oxidatie, hydratatie, oxidatie (dehydrogenatie) en thiolyse. Zie de figuur hieronder.
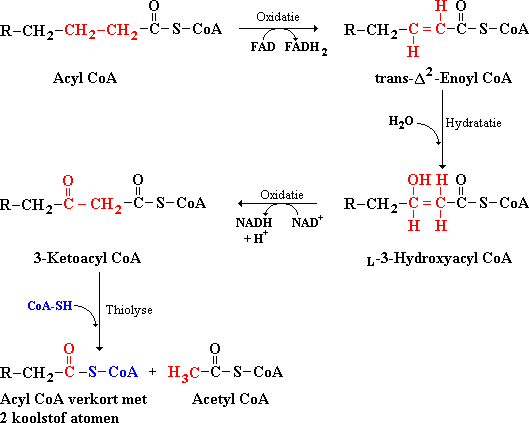
Figuur: Reactie volgorde voor de afbraak van vetzuren: oxidatie, hydratatie, oxidatie en thiolyse.
De drie reacties vanaf acyl CoA t/m 3-ketoacyl CoA zijn vergelijkbaar met de reacties van barnsteenzuur t/m oxaalacetaat in de citroenzuurcyclus.
De afbraak van vetzuren met een keten van een oneven aantal koolstofatomen leidt tot de vorming van propionyl CoA in de laatste thiolyse reactiestap. In de laatste reactiestap van de vetzuurafbraak wordt 3-ketopentanoyl CoA (5 koolstofatomen) gesplitst in propionyl CoA (3 koolstofatomen) en acetyl CoA (2 koolstofatomen). Propionyl CoA wordt omgezet in succinyl CoA (barsteenzuur CoA) en in de citroenzuurcyclus verder afgebroken.
Voor de oxidatie van onverzadigde vetzuren zijn nog een isomerase en een reductase nodig
De eerste reactie in de cyclus van de afbraak (β-oxidatie) van een vetzuur is de oxidatie van een acyl CoA onder vorming van een enoyl CoA met een trans dubbele binding tussen koolstof nummer 2 en 3, zie de figuur hierboven.
Bij de afbraak van een onverzadigd vetzuur verhindert de aanwezigheid van een dubbele binding tussen C-3 en C-4 de vorming van een trans dubbele binding tussen C-2 en C-3. Een trans dubbele binding is nodig voor de vorming van L-3-hydroxyacyl CoA, omdat hiervoor het dehydrogenase enzym specifiek is. Een isomerase verandert een dubbele binding tussen C-3 en C-4 in een trans dubbele binding tussen C-2 en C-3.
Bij de afbraak van een meervoudig onverzadigd vetzuur vormt een cis-Δ4 dubbele binding een ander probleem. Door dehydrogenering van dit deeltje ontstaat een 2,4-dienoyl tussenproduct, dat geen substraat is voor het volgende enzym in de β-oxidatie. Dit probleem wordt omzeild door het enzym 2,4-dienoyl CoA reductase dat met NADPH als coenzym het tussenproduct reduceert tot een cis-Δ3-enoyl CoA. Het eerder genoemde isomerase zet cis-Δ3-enoyl CoA om in de trans vorm, zie de figuur hieronder.
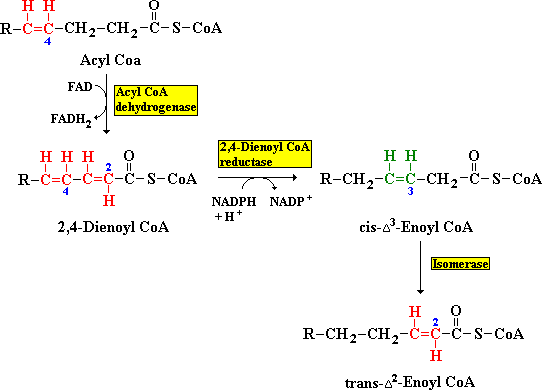
Figuur: Twee enzymen (Acyl CoA dehydrogenase en 2,4-Dienoyl CoA reductase) maken het mogelijk dat onverzadigde vetzuren met een dubbele band op een even koolstofatoom kunnen worden afgebroken.
Oneven genummerde dubbele bindingen worden verwerkt door het isomerase en even genummerde dubbele bindingen door de combinatie van het reductase en het isomerase.
Als de vetafbraak overheerst worden uit acetyl CoA ketonstoffen gevormd
Alle bij de vetzuurafbraak gevormde actief azijnzuur (acetyl CoA) kan alleen voldoende snel in de citroenzuurcyclus verder worden afgebroken wanneer er voldoende oxaalacetaat (oxaalazijnzuur) aanwezig is. Bij vasten of bij diabetes (suikerziekte) wordt oxaalacetaat gebruikt voor de gluconeogenese. Er is dan onvoldoende oxaalacetaat beschikbaar om met acetyl CoA te reageren.
Onder deze omstandigheden vormen twee moleculen acetyl CoA één acetoacetyl CoA en worden daaruit de ketonstoffen gevormd: acetylacetaat (diaceet), D-3-hydroxybutyraat en aceton.
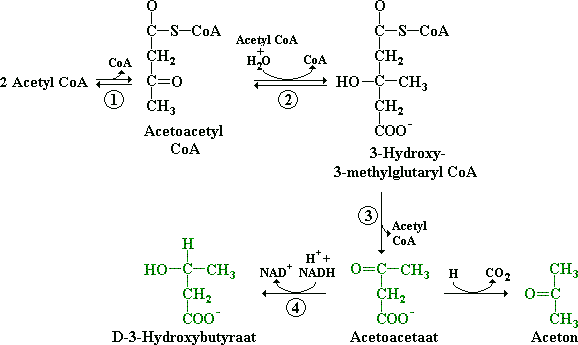
Figuur: Twee moleculen acetyl CoA vormen één acetoacetyl CoA en hieruit worden de ketonstoffen gevormd: acetylacetaat, D-3-hydroxybutyraat en aceton.
De enzymen die deze reacties in de lever versnellen zijn (1) 3-ketothiolase, (2) hydroxymethylglutaryl CoA synthetase, (3) hydroxymethylglutaryl CoA splitsingsenzym en (4) het mitochondriële enzym D-3-hydroxybutyraatdehydrogenase. Acetylacetaat decarboxyleert (=koolstofatoom er vanaf) spontaan tot van aceton. Aceton is een vluchtige stof en men ruikt de geur van aceton in de adem van mensen met diabetes of bij mensen die vasten.
Acetylacetaat is een belangrijke brandstof in sommige weefsels
De ketonstoffen blijken belangrijke energiebronnen te zijn, het is de primaire brandstoffen voor de hartspier en de nierschors. Bij vasten of diabetes schakelen de hersenen van glucose over op het gebruik van acetylacetaat als brandstof.
Acetylacetaat wordt geactiveerd door de overdracht van het CoA van succinyl CoA naar acetylacetaat. Acetoacetyl CoA wordt dan gethiolyseerd tot twee moleculen acetyl CoA die de citroenzuurcyclus ingaan.
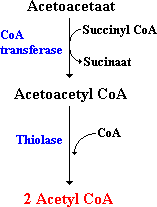
Figuur: Het gebruik van Acetoacetaat als brandstof. Acetoacetaat wordt omgezet in 2 moleculen acetyl CoA, wat de citroenzuurcyclus in kan.
De lever kan acetylacetaat (niet gethiolyseerd) leveren aan andere organen omdat de lever zelf het enzym CoA transferase niet heeft. Andere weefsels hebben dit enzym wel. Acetylacetaat heeft een regulerende rol. Hoge concentraties in het bloed zijn een signaal voor een overmaat aan acetyl-eenheden en leiden tot een vertraagde lipolyse (vetafbraak) in vetweefsel (negatieve feedback).
Mens en dier kunnen vetzuren niet in glucose omzetten. Mens en dier kunnen vetzuren niet in glucose omzetten omdat zij het acetyl CoA niet kunnen gebruiken om pyruvaat of oxaalacetaat te maken. De beide koolstofatomen worden wel in de citroenzuurcyclus opgenomen, maar door twee decarboxylaties wordt per saldo geen extra oxaalacetaat (geen gluconeogenese) gevormd. Planten kunnen dat met behulp van de glyoxylaat cyclus wel.
Vet opbouw (Vetzuur synthese)
Kenmerkende verschillen tussen de afbraak en synthese van vetzuren.
| Afbraak van vetzuren | Opbouw van vetzuren | |
|---|---|---|
| In welk deel van de cel | mitochondriën | cytoplasma |
| Binding van tussenproducten aan | coenzym A | acyl transporteiwit ACP |
| Enzymsysteem | afzonderlijke enzymen | enzymen in één eiwitketen |
| Verandering van de ketenlengte | afsplitsing van C2 (acetyl CoA) | toevoeging van C2 donor: malonyl ACP |
| Redox | oxidatoren: FAD en NAD+ | reductor: NADPH |
De ketenverlenging houdt op na de vorming van palmitaat (C16). Een verdere ketenverlenging en het invoegen van dubbele bindingen worden gekatalyseerd door andere enzymsystemen en vindt plaats in de peroxisomen.
De vorming van malonyl coenzym A is de snelheidsbepalende stap in de vetzuursynthese
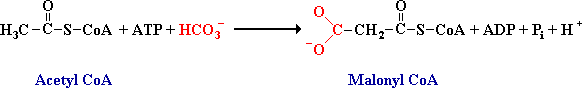
De vetzuursynthese begint met de carboxylering van acetyl Coa tot malonyl CoA door het enzym acetyl CoA carboxylase met biotine als hulp stof. De carboxylgroep van het gevormde malonyl CoA is afkomstig van een bicarbonaation. Deze irreversibele reactie bepaalt de snelheid van de vetzuursynthese. De reactie is vergelijkbaar met de carboxylering van pyruvaat (gluconeogenese).
De cyclus van de ketenverlenging in de vetzuursynthese
Het enzymsysteem dat de opbouw van verzadigde lange-keten vetzuren uit acetyl CoA, malonyl CoA en NADPH katalyseert wordt het vetzuursynthase genoemd. De enzymen van het vetzuursynthase raken los van elkaar wanneer de cellen worden opengebroken. De beschikbaarheid van deze losse enzymen hebben de opheldering van de stappen in de vetzuursynthese vergemakkelijkt.
De tussenproducten in de vetzuursynthese worden gebonden aan een acyl transport eiwit (ACP= acyl carrier protein). De verlengingsfase van de vetzuursynthese begint met de vorming van acetyl-ACP en malonyl-ACP door respectievelijk acetyl transacylase en malonyl transacylase:
Acetyl CoA + ACP ![]() Acetyl-ACP + CoA
Acetyl-ACP + CoA
Malonyl CoA + ACP ![]() Malonyl-ACP + CoA
Malonyl-ACP + CoA
De synthese van vetzuren met een oneven aantal koolstofatomen begint met de vorming van propionyl-ACP uit propionyl CoA door het minder specifieke enzym acetyl transacylase.
De vier reacties van de ketenverlenging in de vetzuursynthese zijn een condensatie, reductie, dehydratatie en een reductie.
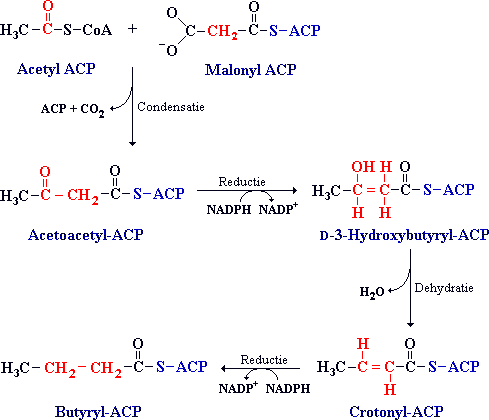
De getoonde tussenproducten worden in de eerste ronde van de synthese gevormd.
Uitleg bij de vier reacties
Acetyl-ACP en malonyl-ACP condenseren tot acetoacetyl-ACP onder invloed van het enzym acyl-malonyl-ACP condenserend enzym. In deze condensatiereactie wordt een C4-eenheid gevormd uit een C2- en een C3-eenheid terwijl er een CO2-groep wordt afgesplitst.
Dat niet twee moleculen acetyl-ACP condenseren heeft als achtergrond dat de reactie met malonyl-ACP een grotere evenwichtsconstante heeft omdat de decarboxylering een belangrijke bijdrage aan de verlaging van de vrije energie levert. De vrije energie die bij de carboxylering van acetyl CoA door ATP in malonyl CoA is vastgelegd komt bij de decarboxylering vrij. Bij de decarboxylering wordt hetzelfde koolstofatoom afgesplitst als dat bij de carboxylering van acetyl CoA door het bicarbonaation is geleverd. Dus, de koolstofatomen van vetzuren met een even aantal koolstofatomen zijn allemaal afkomstig van acetyl CoA.
In de drie volgende reacties van de vetzuursynthese wordt de ketogroep (C met dubbel gebonden zuurstof) aan het derde koolstofatoom gereduceerd tot een methyleengroep (-CH2-). In de eerste reactie wordt acetoacetyl-ACP gereduceerd tot D-3-hydroxybutyryl-ACP. Deze reactie verschilt in twee opzichten van de overeenkomstige reactie in de vetzuurafbraak:
(1) In plaats van het L-epimeer wordt het D-epimeer gevormd.
(2) NADPH is het reducerend middel, terwijl NAD+ het oxiderend middel in de Beta-oxidatie is. Dit verschil is een voorbeeld van het algemene principe dat NADPH gebruikt wordt voor synthetische (opbouw) reacties terwijl NADH wordt gemaakt in energieleverende reacties. Vervolgens wordt D-3-hydroxybutyryl-ACP gedehydrateerd tot crotonyl-ACP. In de laatste stap wordt crotonyl-ACP gereduceerd tot butyryl-ACP, waarmee de eerste verlengingscyclus is voltooid.
Na de eerste ronde is er butyryl-ACP gevormd. In de tweede ronde condenseert butyryl-ACP met malonyl-ACP. Hierna gaan de reacties zoals in de eerste ronde. Zo worden er elke ronde 2 koolstofatomen aan toegevoegd. Dit gaat door tot er palmitaat (C16) is gevormd. Palmitaat wordt dan van ACP afgehaald en er ontstaat palmitaat en ACP.
De nettoreactie voor de synthese van palmitaat is:
8 acetyl CoA + 7 ATP + 14 NADPH + 6 H+ ![]() palmitaat + 14 NADP+ + 8 CoA + 6 H2O + 7 ADP + 7 Pi
palmitaat + 14 NADP+ + 8 CoA + 6 H2O + 7 ADP + 7 Pi
Citraat transporteert acetylgroepen uit de mitochondriën naar het cytoplasma voor de vetzuursynthese
Vetzuren worden uit acetyl CoA gesynthetiseerd in het cytoplasma terwijl acetyl CoA wordt gevormd uit pyruvaat in de mitochondriën.
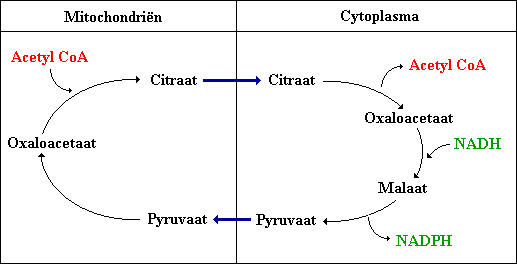
Acetyl CoA wordt uit de mitochondriën naar het cytoplasma getransporteerd in de vorm van citraat (citroenzuur). Tegelijkertijd wordt NADH omgeruild voor van NADPH bij deze reeks van reacties.
Herkomst van NADPH voor de vetzuursynthese
Voor elk acetyl CoA dat wordt verplaatst van het mitochondrium naar het cytoplasma wordt een NADPH gegenereerd. Hieruit valt in te zien dat bij de vorming van palmitaat acht NADPH worden gevormd als gevolg van het transport van acht moleculen acetyl CoA naar het cytoplasma. De overige zes benodigde NADPH komen uit het pentose fosfaat pad.
Bij de regulering van de vetzuurstofwisseling speelt acetyl CoA een sleutelrol
De vetzuurstofwisseling wordt zodanig gereguleerd dat de vorming en afbraak van vetzuren sterk reageren op de behoeften aan energie en andere stoffen. Zo is de vetzuursynthese maximaal als er koolhydraten en energie voldoende zijn en vetzuren schaars.
De snelheidsbepalende stap in de vetzuursynthese wordt gekatalyseerd door acetyl CoA carboxylase. Het enzym wordt gereguleerd door de hormonen adrenaline, insuline en glucagon. Deze boodschapperstoffen signaleren de totale behoeften van het organisme. Insuline activeert de vetzuursynthese. Glucagon en adrenaline hebben het tegengestelde effect.
De regulering wordt ook uitgeoefend door de concentraties in de cel van citraat, palmitoyl CoA en AMP. Het sleutelenzym reageert dus op aan het organisme betreffend regulering en aan locale regulering.
Acetyl CoA carboxylase wordt geïnactiveerd als gevolg van fosforylering door een AMP afhankelijk proteïne kinase. Het wordt geactiveerd door de binding van citraat.